Les membranes cellulaires eucaryotes sont composées de plusieurs milliers de structures de lipides différentes. Pourquoi les cellules déploieraient-elles un tel répertoire si ce n’était que pour conférer aux lipides le seul rôle de barrière entre les milieux extérieurs et intérieurs ? Précisément, les lipides sont impliqués dans de nombreux autres processus cellulaires essentiels, par exemple la signalisation cellulaire, le trafic membranaire, ou la régulation de l’activité des protéines membranaires. Au-delà de la diversité, les membranes cellulaires sont caractérisées par une distribution asymétrique des lipides qui les composent leurs deux feuillets. Le maintien et la dissipation de l’asymétrie des membranes est régulé par trois types de transporteurs (Figure 1) : les flippases (transport du feuillet exoplasmique au feuillet cytosolique), les floppases (transport du feuillet cytosolique au feuillet exoplasmique) et les scramblases (transport bidirectionnel).
Le rôle essentiel du maintien de l’asymétrie des lipides au sein des membranes cellulaires est illustré par le fait que des mutations de flippases (ou ATPases-P4) conduisent chez l’humain à diverses pathologies, par exemple des troubles neurologiques sévères ou une maladie hépatique rare, la cholestase intrahépatique familiale progressive. Par ailleurs, des flippases de la levure pathogène
Figure 1: Les transporteurs de lipides (adapté de Montigny et al, 2016). Gauche, Les flippases transportent des lipides du feuillet exoplasmique (out) au feuillet cytosolique des membranes alors que les floppases catalysent un transport dans la direction opposée. Les scramblases rompent l’asymétrie de sphospholipides par un transport de lipides rapide, bi-directionnel, spontané et peu spécifique. La flèche indique la directionnalité du transport. Les flippases et les floppases catalysent un transport actif primaire. Droite, Topologie des ATPases-P4 et de leus sous-unités associées, les protéines Cdc50. Le domaine transmembranaire est coloré en marron, le domaine de déphosphorylation (A) en jaune, le domaine de fixation du nucléotide (N) en rouge, et le domaine de phosphorylation en bleu. Cdc50, avec deux segments transmembranaires et une grand boucle exoplasmique, est représentée en rose. Les pont disulfures et les sites de glycosylation sont indiqués.
Au laboratoire, nous souhaitons comprendre le mécanisme moléculaire du transport de lipides catalysé par les flippases, ainsi que le lien entre remodelage des membranes par les flippases et biogenèse des vésicules membranaires. À ces fins, nous avons mis au point un système pour la co-expression et la purification de la flippase de la levure S. cerevisiae Drs2/Cdc50 (Jacquot et al, 2012 ; Azouaoui et al, 2014 ; Azouaoui et al, 2016), et identifié la phoshatidylsérine (PS) et le phosphatidylinositol-4-phosphate (PI4P) comme ligands essentiels pour la stabilité du complexe purifié (Azouaoui et al, 2014). Nous avons par ailleurs montré que les extrémités N- et C-terminales de Drs2 jouent un rôle auto-inihibiteur et que le PI4P stimule l’activité du complexe (Azouaoui et al, 2017). L’optimisation de la purification du complexe Drs2/Cdc50 nous a permis de contribuer, en collaboration avec l’équipe de Poul Nissen (Université d’Aarhus, Danemark), à la détermination de la première structure à haute résolution d’une flippase (Timcenko et al, 2019) (Figure 2).
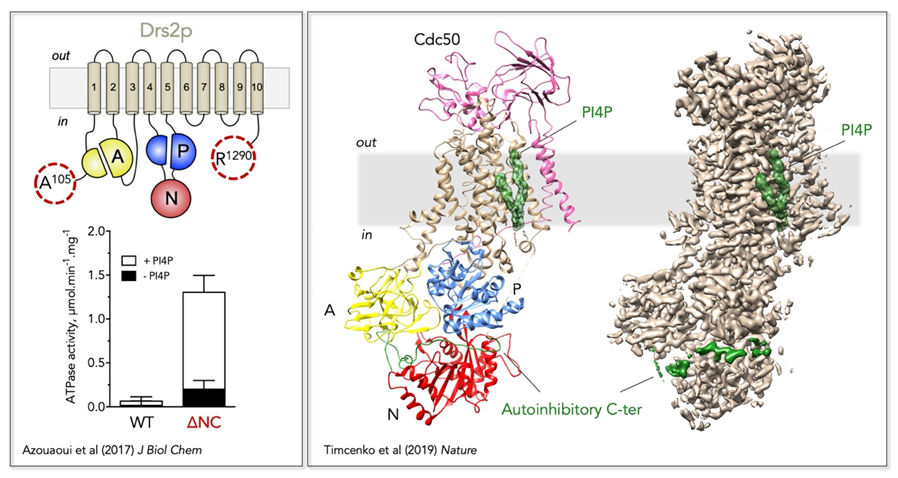
Figure 2. Mécanisme de regulation de la flippase Drs2/Cdc50. Gauche, La coupure protéolytique des extrémités N- et C-terminales de Drs2 stimule son activité et reste sensible à la présence de PI4P. Droite, Localisation des éléments régulateurs de Drs2/Cdc50 sur la structure à haute résolution du complexe (PDB : 6ROI, EMD :4973). Sur le modèle structural (gauche), le domaine transmembranaire est coloré en marron clair, le domaine A en jaune, le domaine N en rouge, et le domaine P en bleu ; le PI4P et le domaine C-terminal auto-inhibiteur sont représentés en vert. Le PI4P et le domaine auto-inhibiteur sont également représentés sur la carte de densité électronique (droite).
Plus précisément, nous développons les projets suivants au laboratoire :
-
Mécanisme de regulation du complexe Drs2/Cdc50 ; même s’il est maintenant connu que le complexe est auto-inhibé par ses ectrémités terminales, quelle partie précisément du C-terminal est directement responsable de l’auto-inhibition et par quel mécanisme reste à élucider. Aussi, quelle est la contribution du N-terminal dans l’auto-inhibition ? De la même manière, bien que le site de fixation du PI4P sur Drs2 ait été identifié (Figure 2), le mécanisme moléculaire de l’activation par le PI4P reste inconnu.
-
Reconstitution in vitro de la machinerie de translocation de lipides. Plusieurs études ont montré une interaction physique entre Drs2 et les protéines Arl1 et Gea2, et que cette interaction stimule le transport de lipides in vivo. Nous cherchons désormais à reconstituer l’assemblage entre Drs2/Cdc50, Arl1 et Gea2
in vitro et à étudier ses propriétés fonctionnelles.
-
Conséquences, au niveau moléculaire, des mutations de la flippase humaine ATP8B1. Certaines mutations d’ATP8B1 conduisent à une pathologie rare, la cholestase intrahépatique progressive familiale. Nous produisons et purifions la flippase ATP8B1 exprimée dans
S. cerevisiae, pour étudier par la suite les conséquences fonctionnelles des mutations d’ATP8B1.
-
Étude de flippases putatives de la levure C. neoformans. L’ATPase-P4 Apt1 est associée à la virulence du pathogène. Ce projet consiste en la caractérisation biochimique et fonctionnelle détaillée de la protéine Apt1 afin, par la suite, d’identifier le potentiel de cette protéine comme cible thérapeutique.
Principales collaborations
- Eva Pebay-Peyroula, Institut de Biologie Structurale, Grenoble
- Bruno Miroux, Institut de Biologie Physico-chimique, Paris
- Joost Holthuis, Université d’Osnabrück, Allemagne
- Poul Nissen et Jesper Vuust Møller, PUMPKIN, Université d’Aarhus, Danemark
- Rosa López-Marqués, PUMPKIN, Université de Copenhague, Danemark
- Isabelle Florent, Muséum d’Histoire Naturelle, Paris
Publications récentes